・初期過程ともいい、光エネルギーを化学エネルギー(ATP&NADPH)に変換する役割を担い、『葉緑体』にあるチラコイド膜上で行われる。
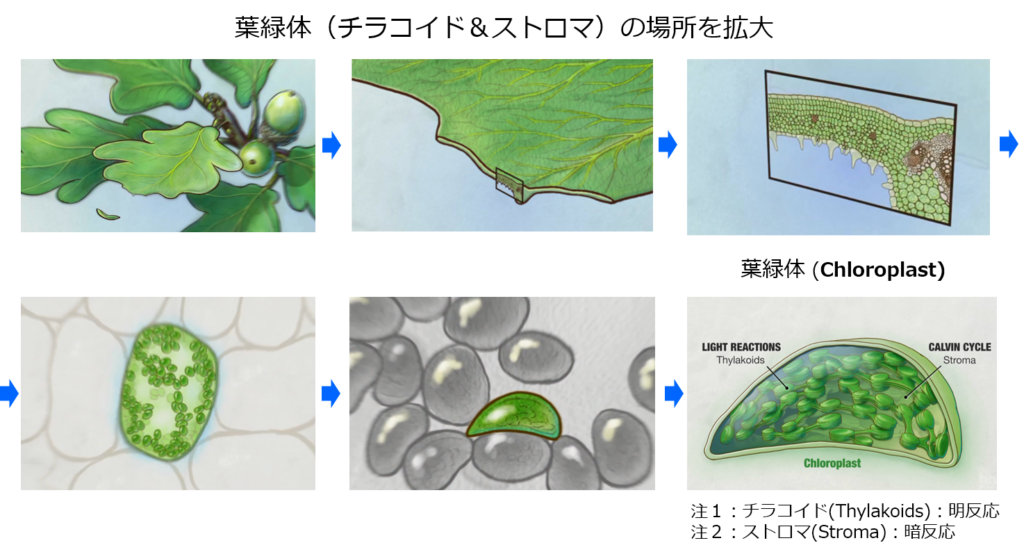
チラコイド膜上では
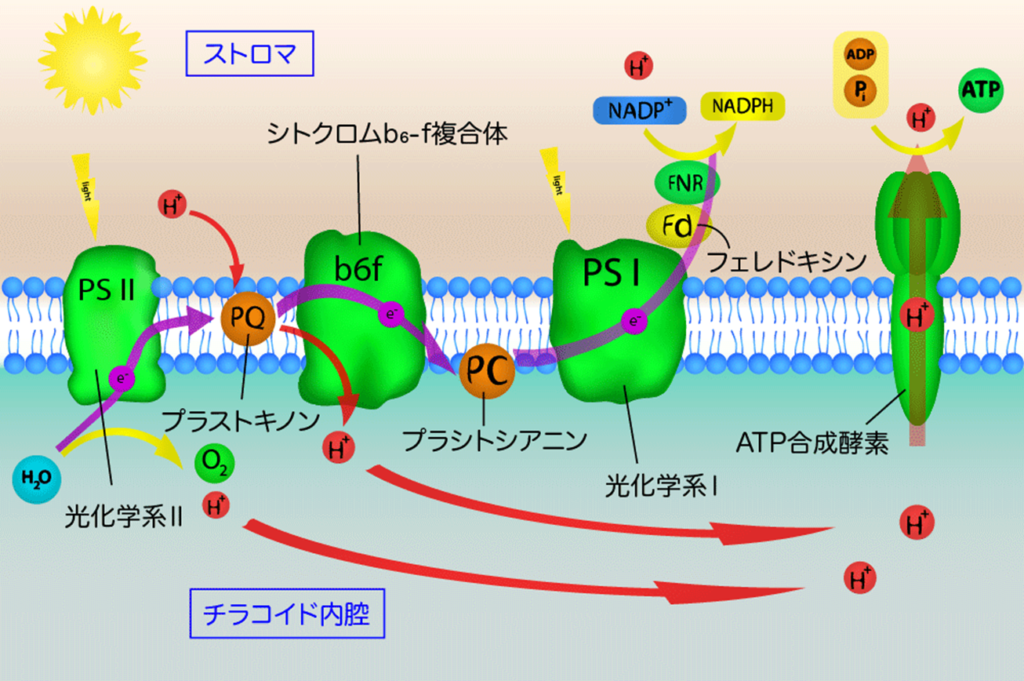
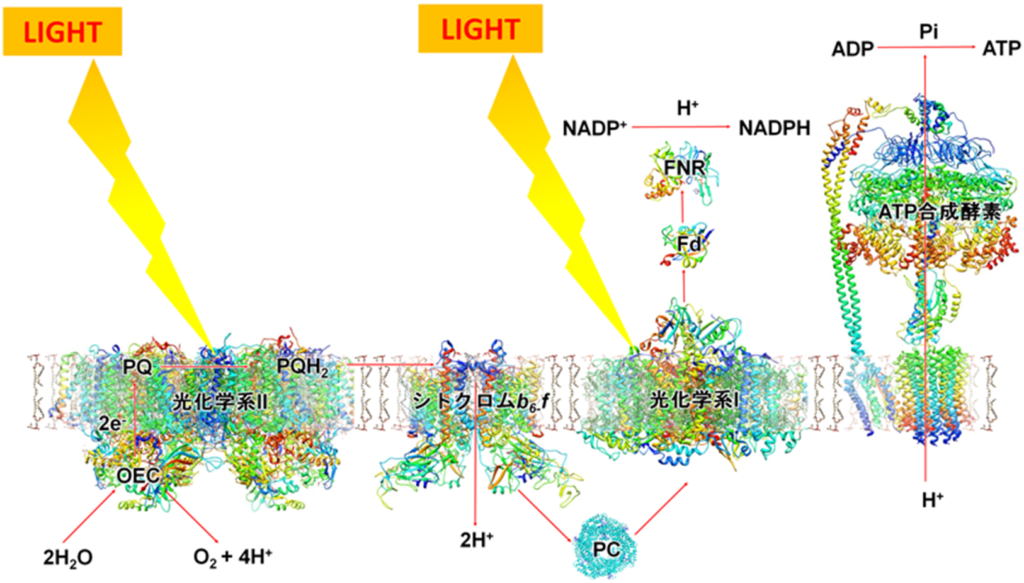
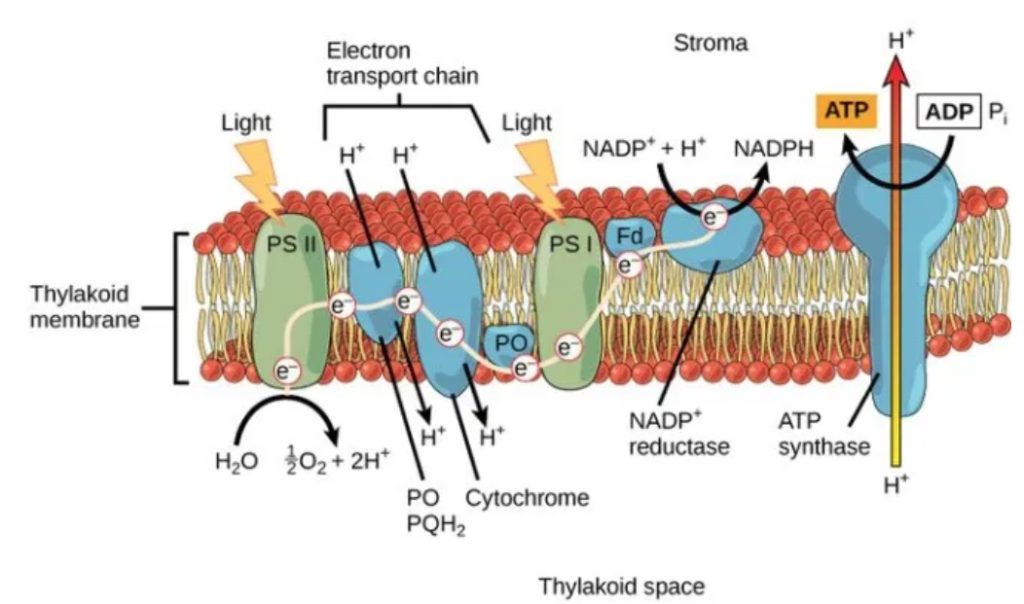
・光化学反応を行う光化学系Ⅱ、Ⅰ(PSⅡ、PSⅠ)が有り、両者間の電子移動(e⁻)はシトクロムb₆/f複合体を介して行われ、最終的には『NADPH』をストラマ側にて生成する。
『光化学系』と呼ばれ、光合成電子伝達反応と定義されている。
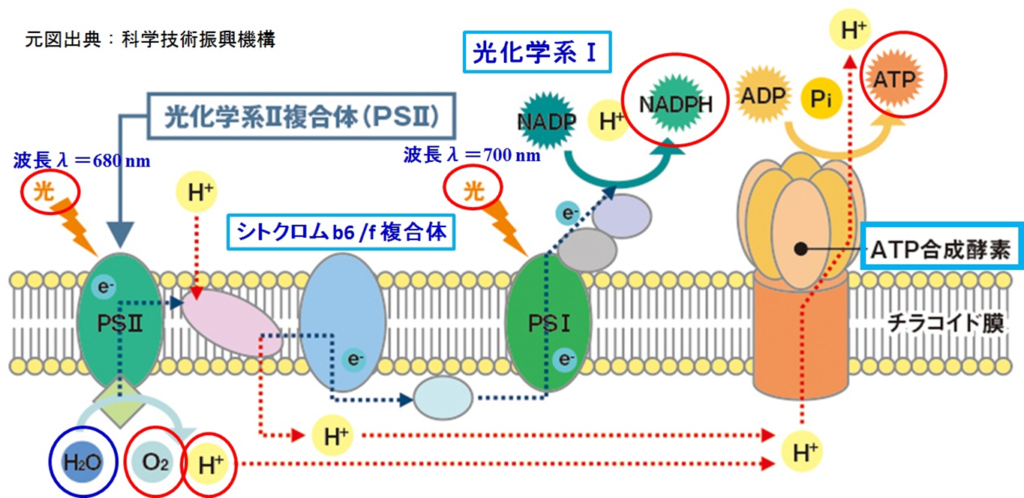
・PSⅡ内で電子(e⁻)と同時に生み出されたH⁺(チラコイド側)及びストラマ側よりH⁺を取り込むプラストキノン(PQ:下図のシトクロムb₆/f複合体の手前:薄紫色の楕円部)はチラコイド側にH⁺を放出し、H⁺濃度勾配(プロトン濃度勾配)を高め、H⁺濃度勾配の解消をさせるATP合成酵の働きで『ATP』をストラマ側にて生成する。
『光リン酸化』と呼ばれ、光合成電子伝達反応と区別して定義されている。
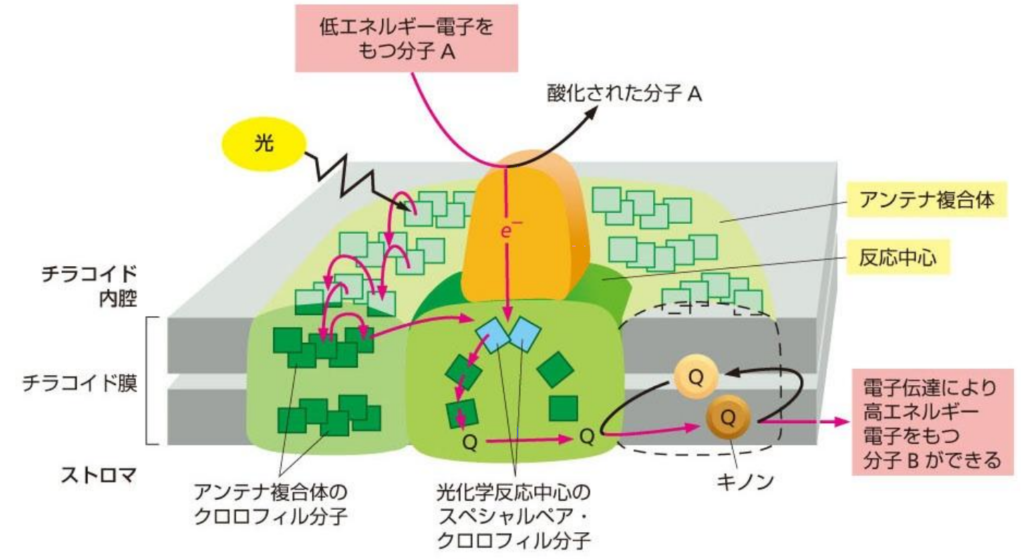
■■明反応である光化学系(PSⅡ、PSⅠ)は更に細分化され、A)集光性複合体とB)反応中心で構成される。
■A:『集光性(Antenna)複合体』
光を捉える色素がタンパク質と結合して集光アンテナを作る。
細胞の分子生物学 第5版
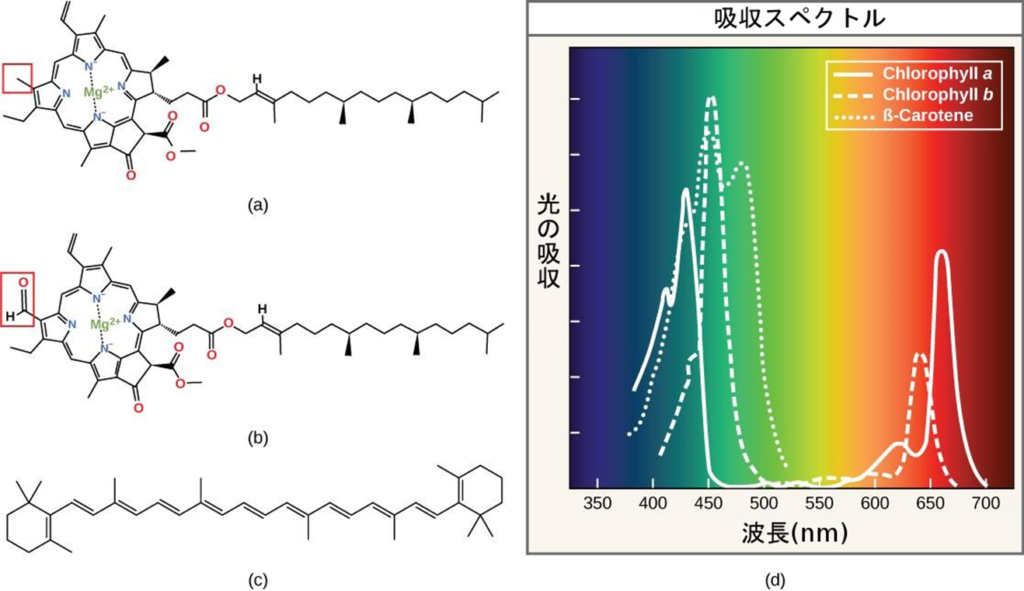
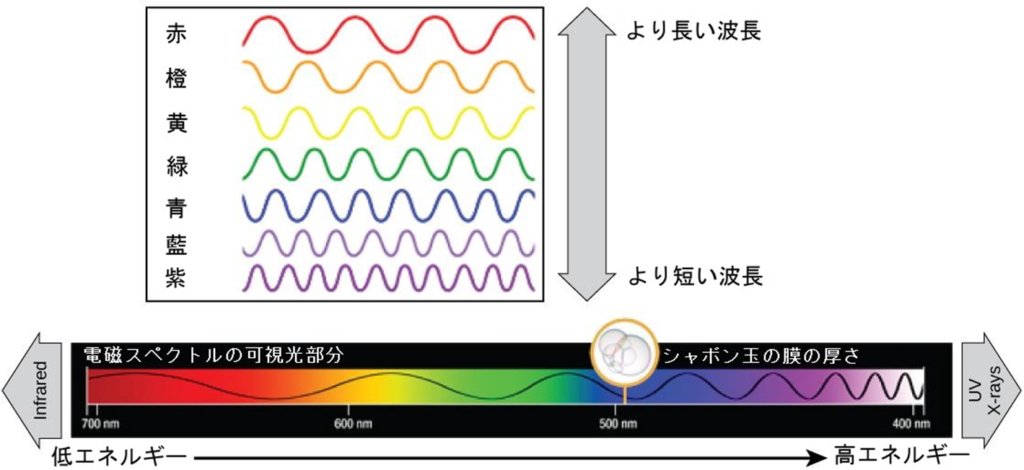
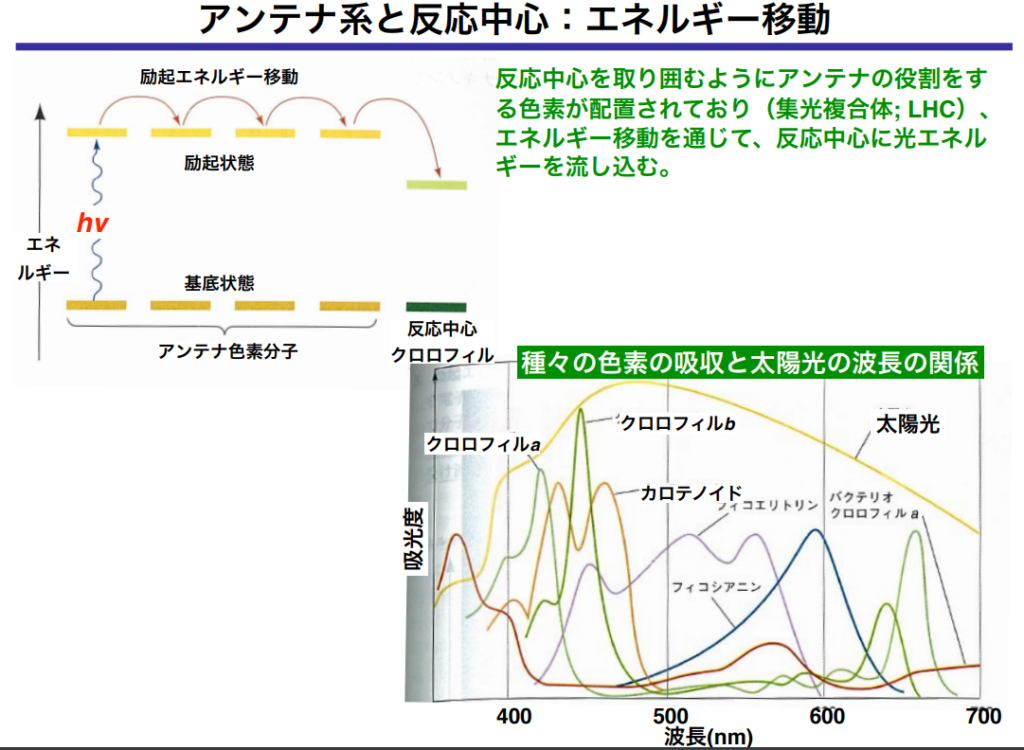
主な色素としてクロロフィルa、クロロフィルb(緑色に見える色素)やβ-カロテン等があり、それぞれのタイプの色素は、可視光から吸収する波長の特定パターンによって識別できる。
尚、光合成に最も効率よく使われる光は青と赤の波長である。
OpenStax のサイトで公開されている教科書“ Biology 2e”
OpenStax のサイトで公開されている教科書“ Biology 2e”
注1)青から紫に向かうほどより短い波長を持ち、それゆえに高いエネルギーを持つ。逆に、黄から赤に向かうほど波長は長くなり、エネルギーは低くなる。(ロープを長くて広い波を動かすにはさほど力を要さないが、短くて狭い波を動かすには大変なエネルギーを要す)
注2)クロロフィルaは可視スペクトルの両端(青と赤)からの波長を吸収するが、緑は吸収しない。緑色が反射または透過するので、クロロフィルは緑色に見える。カロテノイドは、短波長の青色領域を吸収し、より長い波長の黄色、赤色、およびオレンジ色を反射する。
注3)クロロフィルは、中心にMg²⁺を配置し、4つのN(窒素)が取り囲んだ形になっている(出典:生物学 第2版 — 第8章 光合成 —(a)と(b))。Mg(マグネシウム)が光合成において重要な要素になっている。
尚、農業において、一般的に多量要素(植物の要求量の多い肥料養分)に挙げられるのが、窒素(N)、リン酸(P)、カリウム(K)、カルシウム(Ca)、マグネシウム(Mg)である。Mgについては、クロロフィルの中心物資である為に要求量が多い。
Mgが不足すると植物の下葉が黄化する。これは、植物が光合成がより必要となる上位葉にMgを移動させるためである。植物体内ではMgは容易に移動が可能なため、顕著に症状が現れると言われている。

注4)人間の網膜色素は、700nmから400nmの間の波長の光、つまり可視光と呼ばれるスペクトルしか「見る」(吸収する)ことができない。同じ理由で、植物では、色素分子は700nmから400nmの波長範囲の光だけを吸収する。
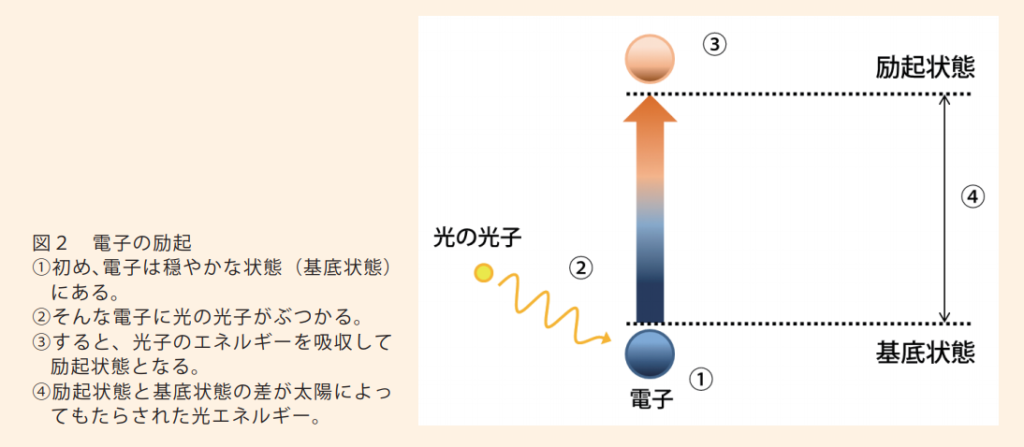
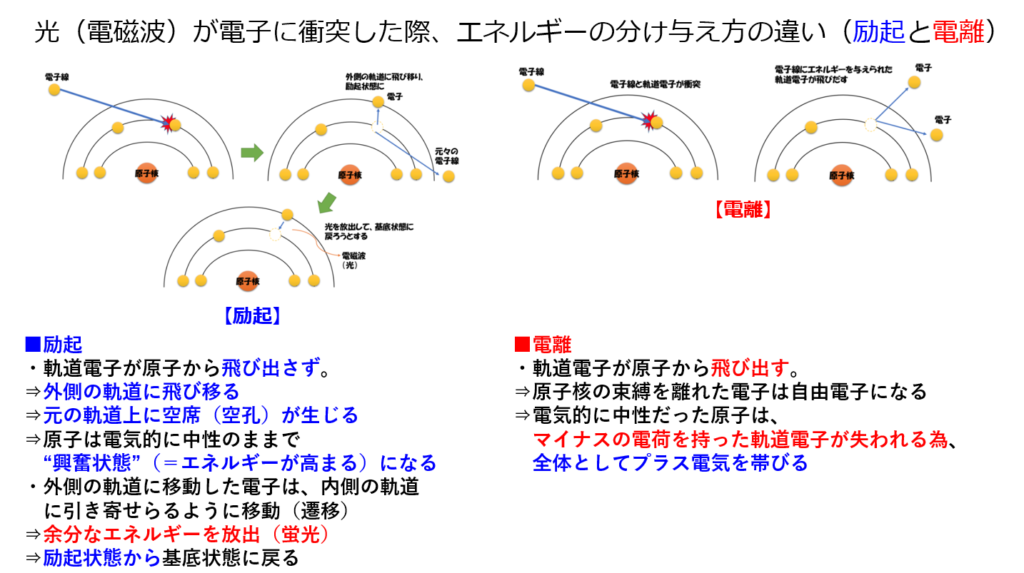
●光が『集光性(Antenna)複合体』に衝突すると
・光子のエネルギーがクロロフィル分子を励起させる。
~燃料電池と生物の仕組みの共通点~
自然農法vol.73 2015.9
http://www.infrc.or.jp/wxp/wp-content/uploads/NFM/NFM_7310.pdf
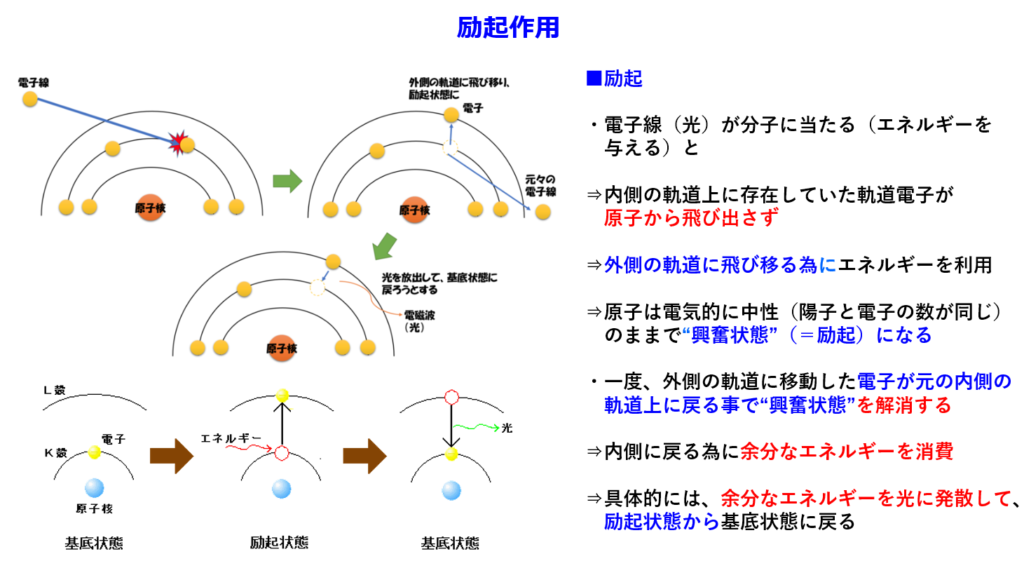
・通常、励起エネルギーは蛍光や熱として発散するが、集光性クロロフィルの場合、吸収されたエネルギーは『共鳴エネルギー移動』によりそのまま隣の分子に伝達される。
細胞の分子生物学 第5版
隣接するクロロフィルを次から次へと励起させ、最終的に反応中心クロロフィル(P680)に効果的に光エネルギーを送り込む。
励起した電子のエネルギーのみを隣のクロロフィルに渡して、基底状態に戻るので、『電子伝達』が起きない。つまり電子の移動ではなく、エネルギーのみが分子間を移動している。
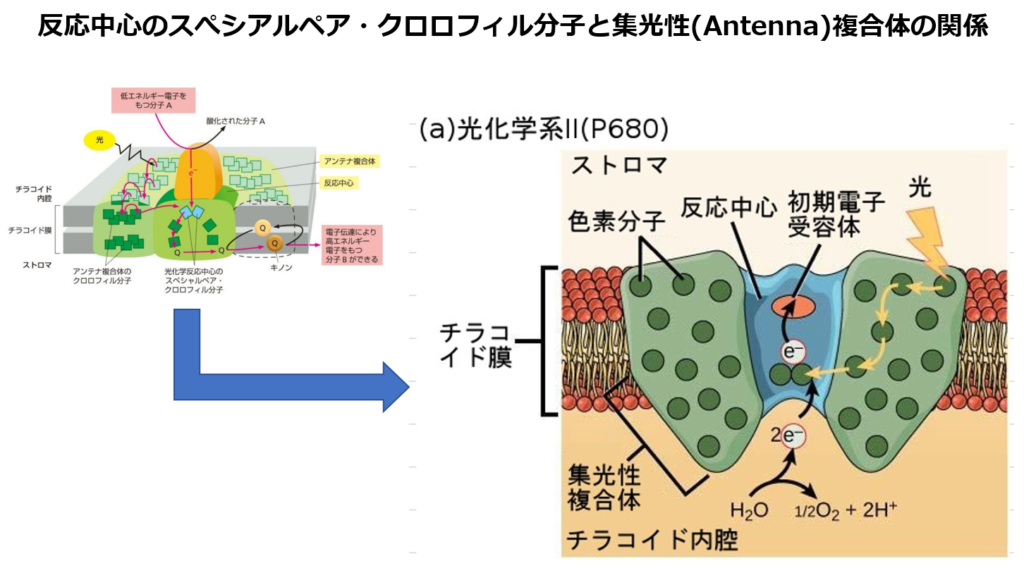
■B:『反応中心』
集光性複合体の中の色素は、反応中心の中の2つの『特別なクロロフィルa対分子=スぺシャルペア・クロロフィル分子』に光エネルギーを渡す。
OpenStax のサイトで公開されている教科書“ Biology 2e”
スぺシャルペア・クロロフィルa対分子は、集めたエネルギーから『電子(e⁻)』を分離させてプラストキノン(PQ)に渡す。
葉緑体の構造と光合成反応を解説
これによって電子移動という反応を開始する。
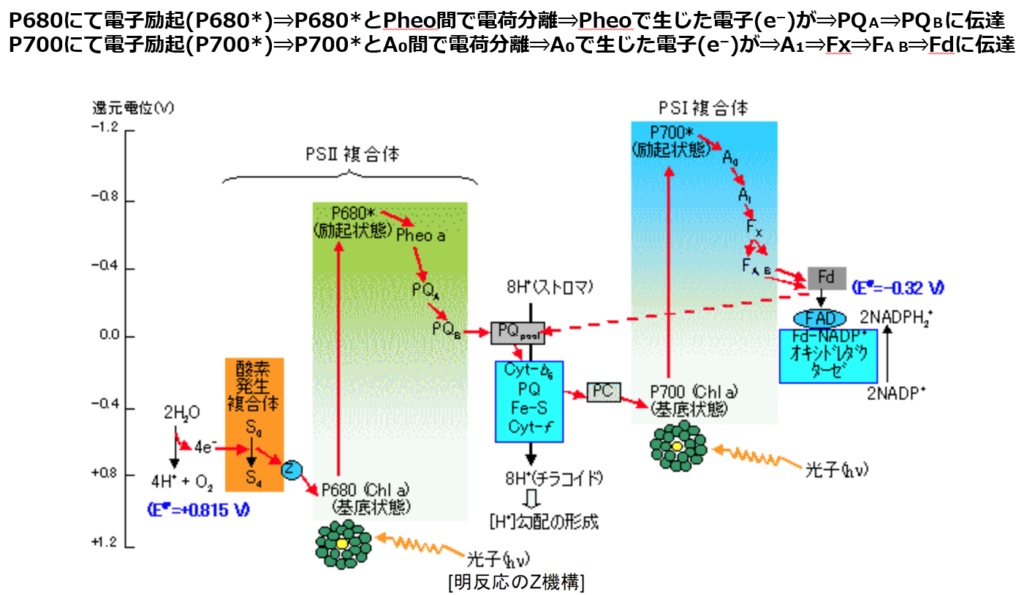
太陽光によって励起されたエネルギーは上図のようにして電子伝達鎖内(PSⅡ→PQ→シトクロムb₆/f複合体→PC→PSⅠ)を移動し、最終的にはNADP⁺にたどり着く。
●『電荷分離反応』を起こす事が出来るスぺシャルペア・クロロフィルa対分子
・電子を放出(電離)する役割を持つ。

・電子が飛び出した後のスぺシャルペア・クロロフィルa対分子(P680)には、どこからか電子を供給されて元の状態に戻らなければ、次の光反応ができない。
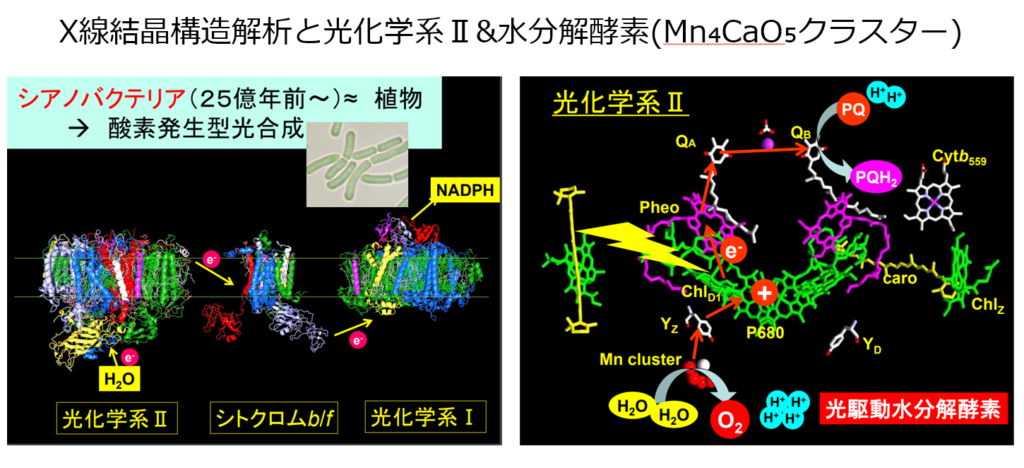
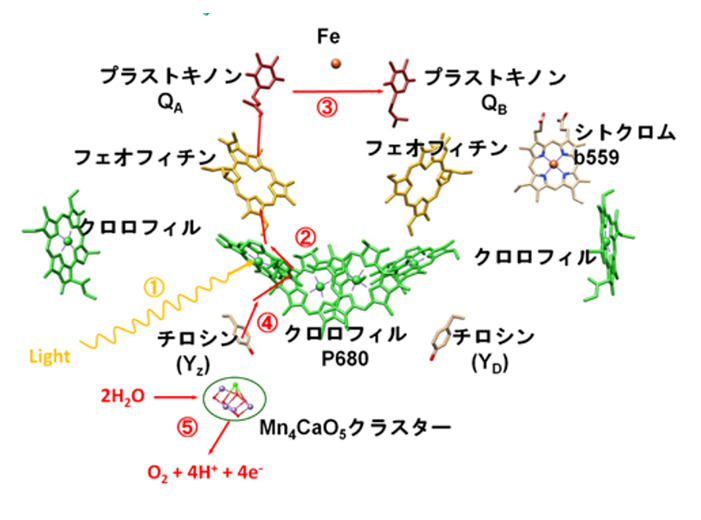
電子供給先は、チロシン残基Yzを経て、Mn₄CaO₅クラスターと呼ばれる金属化合物から電子を奪い取り、元の状態に戻る。
変換と酸素発生の分子機作 G研 野口 巧
https://www.phys.nagoya-u.ac.jp/ex/2010/noguchi.pdf
●Mn₄CaO₅クラスター
Mn₄CaO₅クラスターは『水分子』から電子を奪い取り、2分子の水から4つの電子を奪い取られると、水が分解され、1つの酸素分子を生じる。
水の酸化(酸素の発生)
反応式 2H₂O → O₂ + 4H⁺ + 4e⁻
水は極めて安定的な物資で、通常の光を照射しても分解される事は決してない。
反応式からも分かるように、『水分子』から電子を引き抜く必要がある。
Mn₄Caクラスターは単独では水分解を行う事ができず、必要な酸化力を外部から得る必要がある。
その酸化力を与えるのが、スぺシャルペア・クロロフィルa対分子(P680)である。
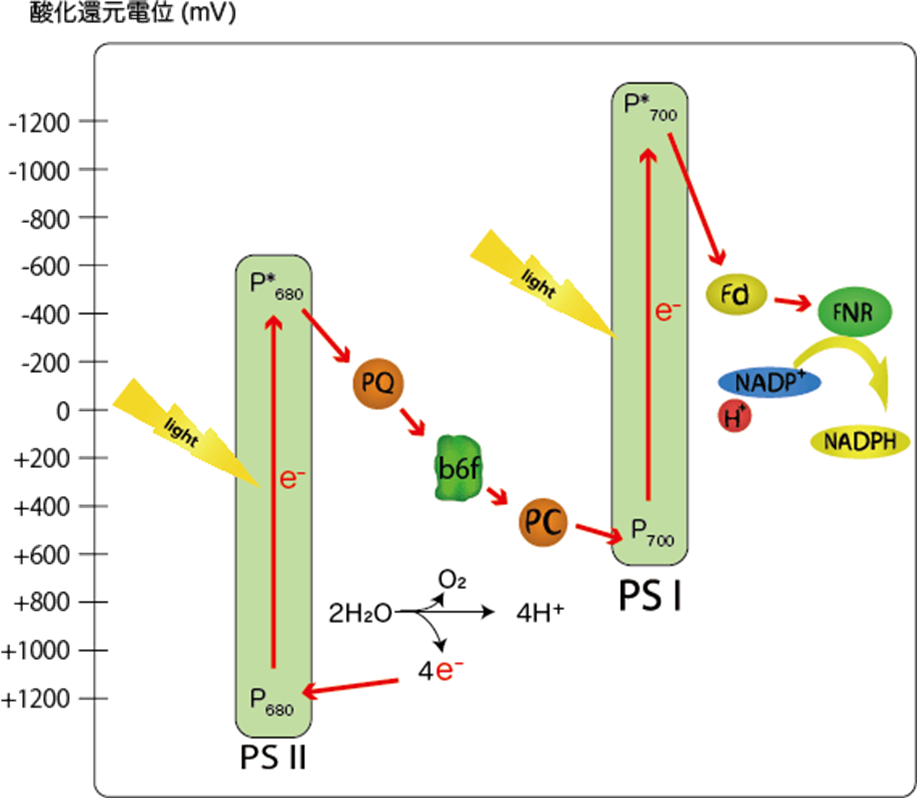
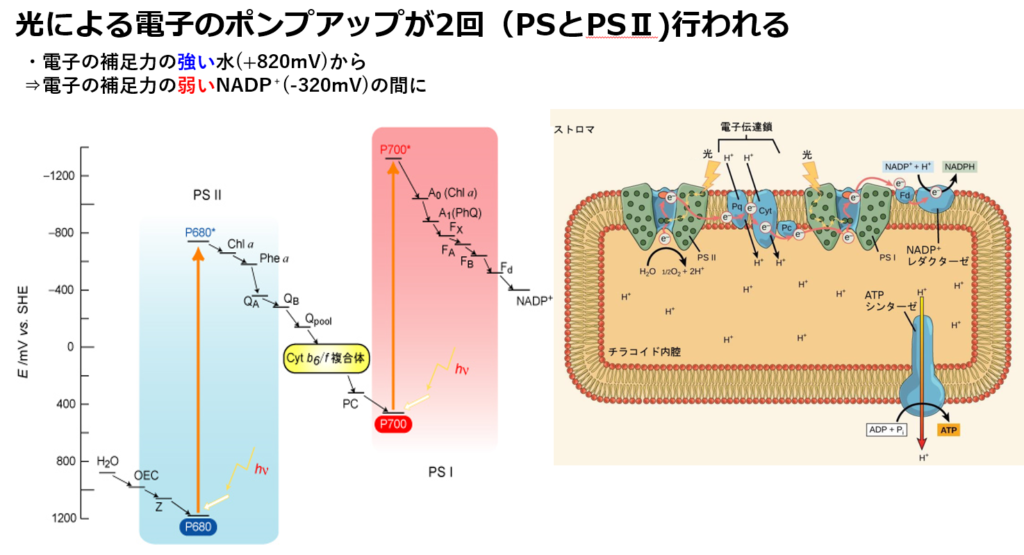
P680は1100~1200mVと異常に高い酸化還元電位を持ち、水(820mV)から電子を引き抜く事が可能である。
葉緑体の構造と光合成反応を解説
注1:電子伝達を行う分子の酸化還元電位の変遷を見ると上図(図1)を横向き(90度右回転)にするとZのように見えるので、Zスキームとも呼ばれる。
注2:この電子伝達系では、酸化還元電位(上図:縦軸)が高い物資ほど電子(e⁻)を保持しやすく、低い物質ほど電子(e⁻)を手放しやすい性質を持つ。
\begin{aligned}\dfrac {1}{2}O_{2}+2H^{+}+2e^{-}\overrightarrow {\leftarrow } \ H_{2}O+820mV\end{aligned}
酸化還元電位が高いという事は、電子を受け取りやすく、手放そうとしない事を意味する。
従って、これより高い酸化還元電位の物資を用意しないと『自ら電子を引き抜く』事ができない。
それを実現するのが
光化学系Ⅱ(PSⅡ)に含まれるスぺシャルペア・クロロフィルa対分子(P680)。
このペアは光の吸収波長を680nmに持つことから、P680と呼ばれる。
その酸化還元電位は+1270mVと非常に強力である。
\begin{aligned}\\p^{+}_{680}+e^{-}\overrightarrow {\leftarrow }P_{680}+1270mV\end{aligned}
結果、光化学系Ⅱ(PSⅡ)では以下の反応が起きる。
2H₂O + 4つの光子 → 4H⁺ + 4e⁻ + O₂
Mn₄CaO₅クラスターから電子を引き抜くステップは5つの中間状態(S₀~S₄)の光駆動サイクルとして『水分解』が行われる。
1 回の光反応 により 1 個の電子が引き抜かれると次の中間状態へと遷移し,計 4 回の電子移動によ り,2 分子の水が酸素とプロトンとに分解 する。
光合成で発生する酸素はこれに由来する。
注:プロトン(H⁺)の放出はS₀→S₁とS₂→S₃では各1個、S₁→S₂では0、S₄→S₀で2個放出される。(4段階から成る反応)
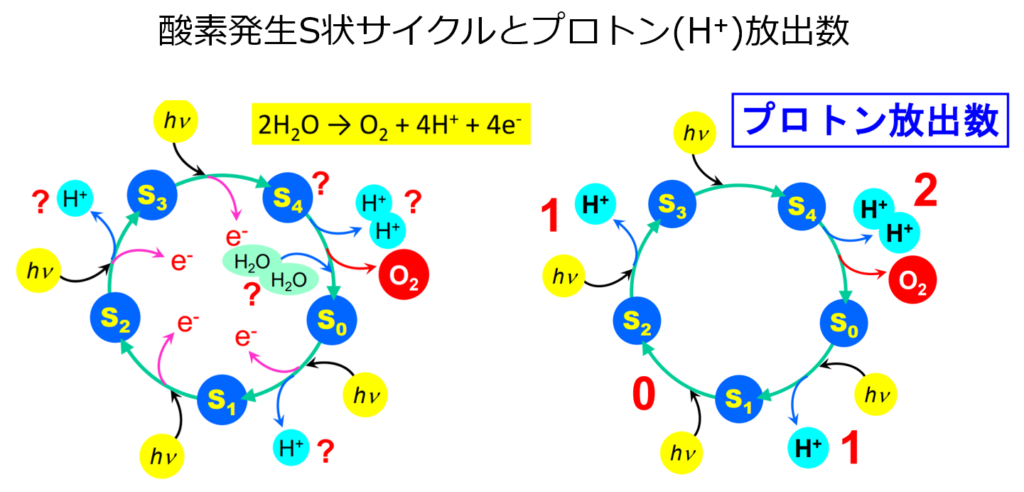
変換と酸素発生の分子機作 G研 野口 巧
https://www.phys.nagoya-u.ac.jp/ex/2010/noguchi.pdf
尚、この反応で生じたH⁺(プロトン)はチラコイド内腔に流入することで濃度勾配形成に利用される。
●SPring-8で解明されたMn₄CaO₅クラスターの構造解析
水分子を分解する触媒(酵素)中心は、4つのマンガン原子(Mn)、1つのカルシウム原子(Ca)、5つの酸素原子(O)、4つの水分子(W)によって構成されている明らかになった。
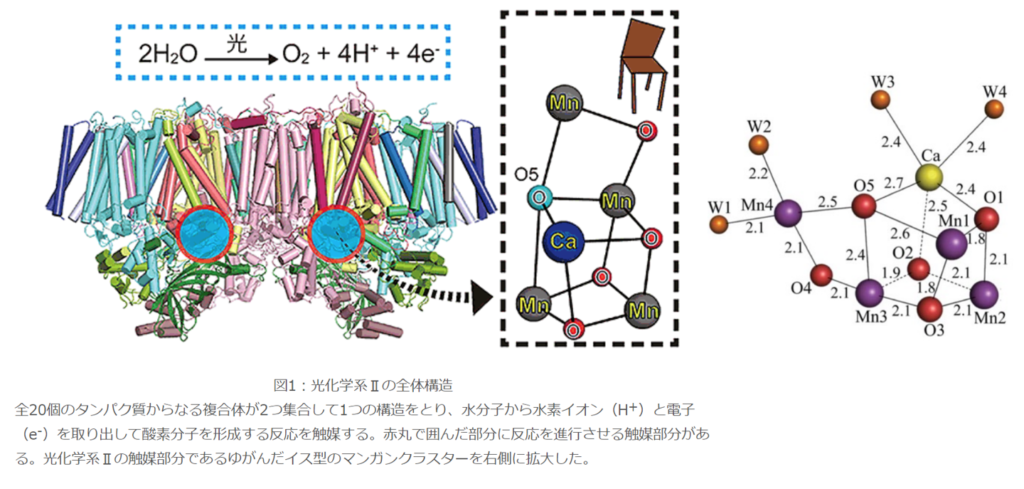
2011年になって初めてMn₄CaO₅クラスターの分子構造が解明され、非対称で、ゆがんだ椅子の形をしている事が判明した。
●電子伝達鎖の経路
PSⅡの反応中心(=スぺシャルペア・クロロフィルa対分子(P680))は励起エネルギーを受け取ると励起され、電子を放出する。この放出された電子が電子伝達系内で伝達が開始される。
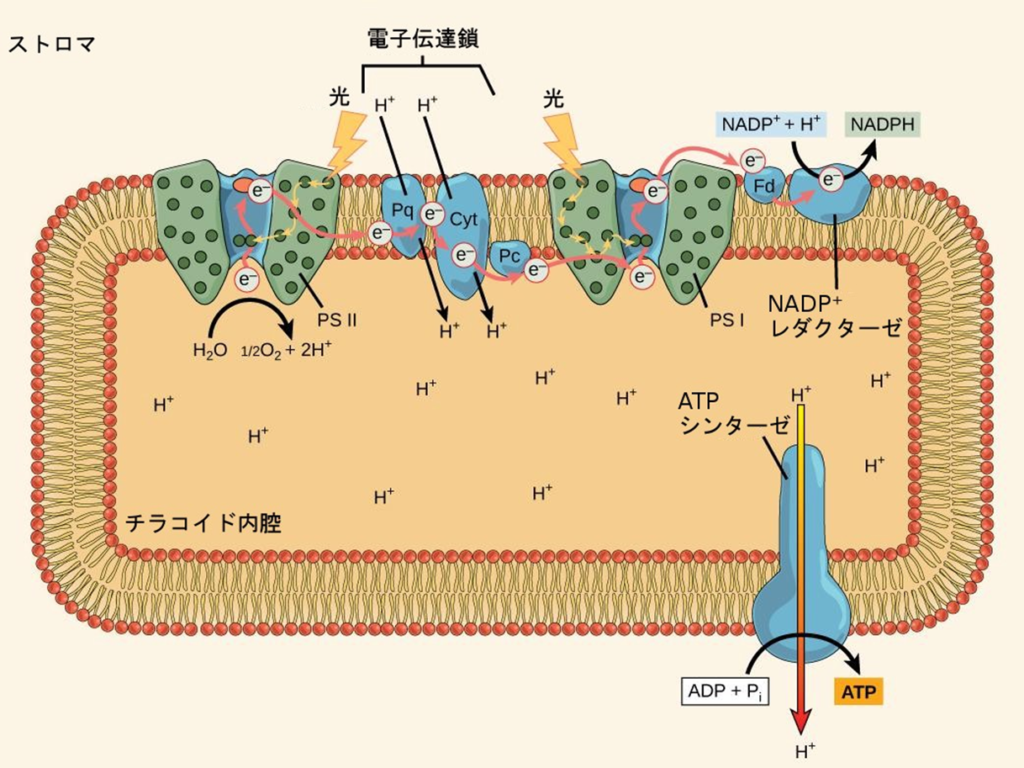
OpenStax のサイトで公開されている教科書“ Biology 2e”
伝達経路は①PSⅡ(P680)⇒②シトクロムb₆/f⇒③PSⅠ(P700)で伝達され、伝達媒介項としてプラストキノール(PQH₂)とプラストシアニン(PC)がある。
①最初の起点は、スぺシャルペア・クロロフィルa対分子(P680)から電子を励起(P680*)する事から始まる。
電子が励起されたP680*とPheo(フェオフィチン)との間で電荷分離が起きる。
www.sc.fukuoka-u.ac.jp › Biochem › photosyn
他方、電子が飛び出した後のスぺシャルペア・クロロフィルa対分子(P680)には、電子(e⁻)を補充する為にチロシン残基Yzから電子(e⁻)を引き抜き、Yzのカチオンラジカルを生成する。ラジカル化したYzはMn₄CaO₅クラスターから電子(e⁻)を引き抜き、Mn₄CaO₅クラスターは強力な酸化力を得る事になる。これによってMn₄CaO₅クラスターは水から電子(e⁻)を引き抜き、プロトン(H⁺)を放出する。(2分子の水から4つの電子(e⁻)が引き抜かれると、4つのプロトン(H⁺)と1つの分子酸素(O₂)が放出される。2H₂O →4e⁻ +4H⁺ + O₂)
秋田 総理,菅 倫寛,沈 建仁
岡山大学異分野基礎科学研究所・光合成・構造生物学研究コア
Pheo(フェオフィチン)で生じた電子(e⁻)がD2 タンパク質に結 合したプラストキノン \(Q_{A}\)に渡され、続いて D1 に結合したプラストキノン \(Q_{B}\) に渡される。
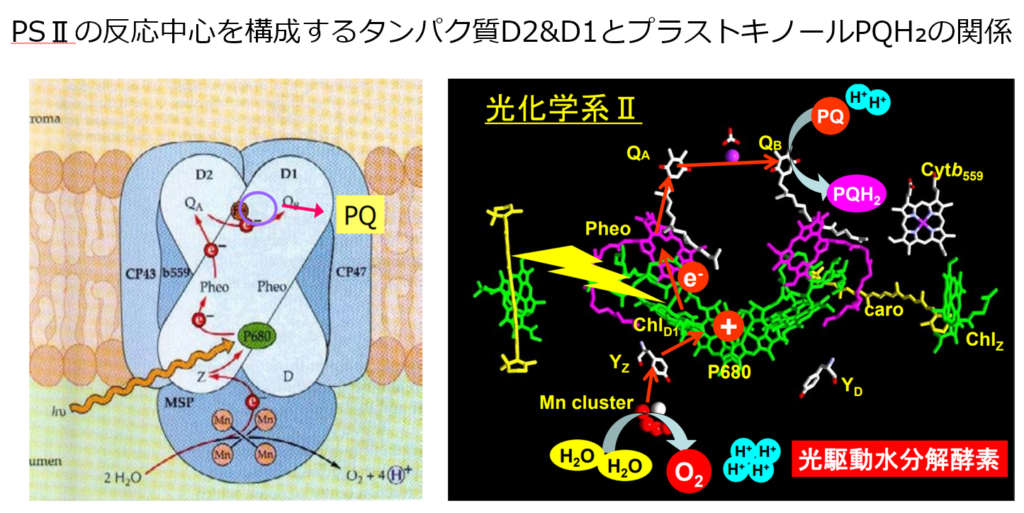
プラストキノン\(Q_{B}\)は2個の電子(e⁻)を受け取るとジアニオンとなり、二つのプロトン(H⁺)をストロマから受け取り、還元型のプラストキノール(PQH₂)になる。
注:2電子(e⁻)を受け取ったキノン分子(PQ)にプロトン(H⁺)が2つ付加すると、ヒドロキノンという安定な物質ができる。

プラストキノール(PQH₂)はPSⅡから遊離したのち、チラコイド膜内を拡散により移動し、『シトクロムb₆/f』へと電子を運ぶ。
尚、プラストキノール(PQH₂)には電子(e⁻)2個とプロトン(H⁺)2個が含まれている。
秋田 総理,菅 倫寛,沈 建仁
岡山大学異分野基礎科学研究所・光合成・構造生物学研究コア
空になった(\(Q_{B}\))結合部位にはプラストキノンプールから新たなプラストキノンが補充される。
②『シトクロムb₆/f』に渡された電子(e⁻)とプロトン(H⁺)の内、プロトン(H⁺)2個はチラコイド内腔に強制的にお送り込まれる。
尚、チラコイド内腔では、2分子の水分解で4つ増えたプロトン(H⁺)と更にシトクロムb₆/fから2個のプロトン(H⁺)が加えられ、益々水素イオン濃度が高くなり、濃度勾配が高くなると表現さてている。
『シトクロムb₆/f』に渡された電子(e⁻)は、酸化型プラストシアニン(PC)を通じてPSⅠに伝達される。
③PSⅠ(P700)は還元型プラストシアニン(PC)から受け取った電子(e⁻)をフェレドキシンに伝達し、還元型フェレドキシンからNADP⁺にに電子伝達を行い、最終還元物質であるNADPHを生産する。
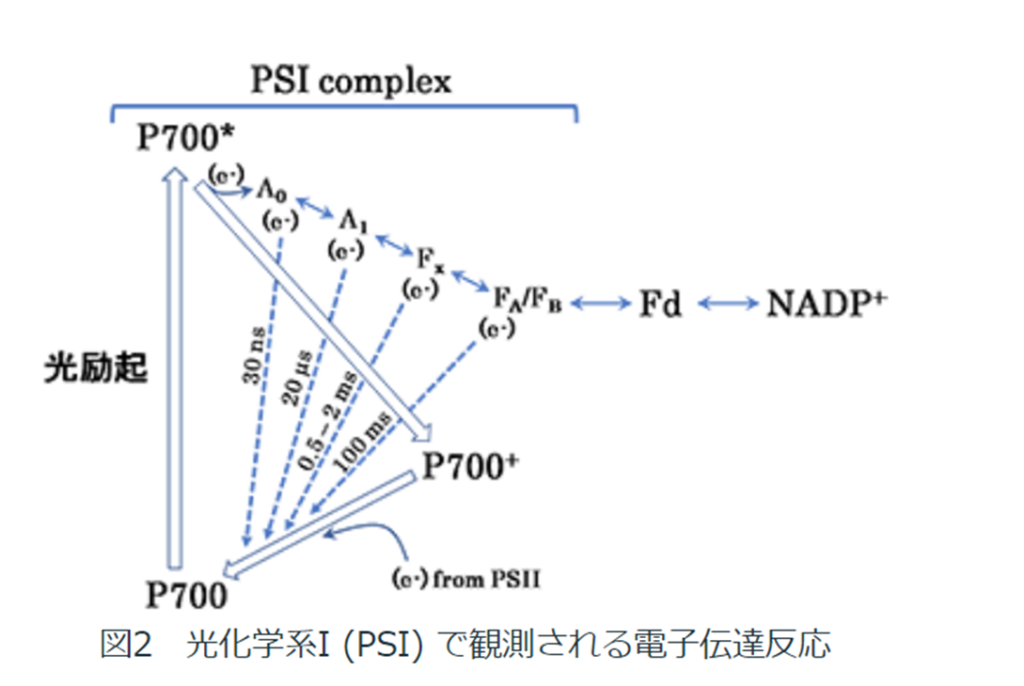
PSⅠの反応段階は以下になる。
1.アンテナ色素タンパク質によって集光された700nmの波長の光でスぺシャルペア・クロロフィルa対分子(P700)が励起される。
電子が励起されたP700*とクロロフイルA₀との間で電荷分離が起きる。
クロロフイルA₀に電子を渡し、P*はP700⁺へ酸化される。
P*はPSⅡから来る電子(e⁻)を受け取り基底状態に還元される。
https://www.kobe-u.ac.jp/research_at_kobe/NEWS/news/2019_06_10_01.html
2.クロロフイルA₀が受け取った電子(e⁻)はA₁、鉄-硫黄クラスター\(F_{X}\)、\(F_{A}\)、\(F_{B}\)を通じて伝達される。
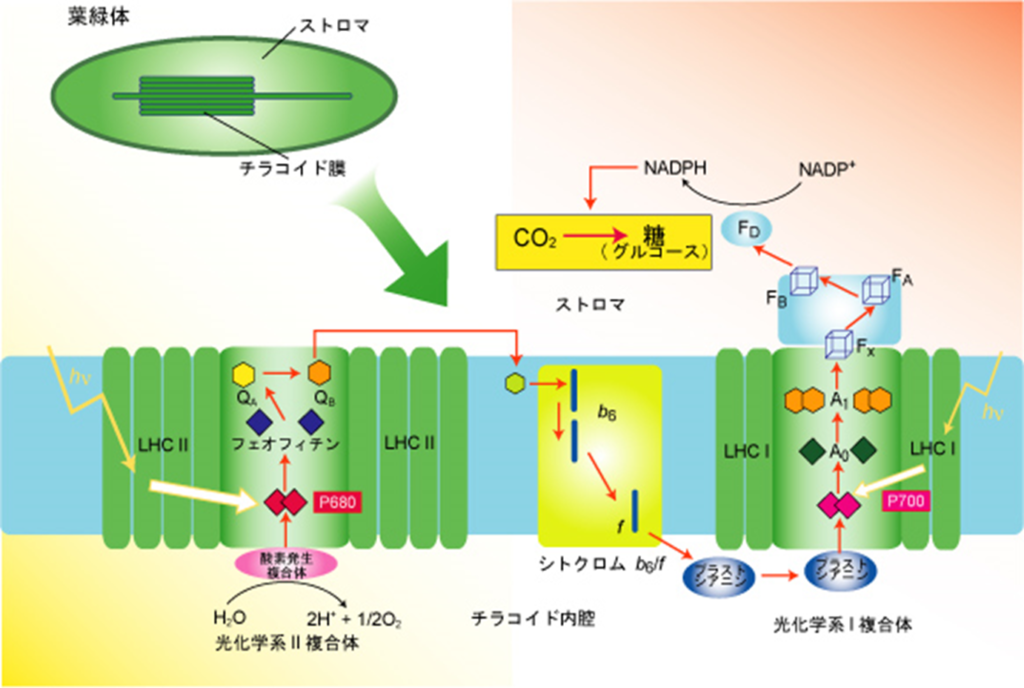
3.還元型フェレドキシン\(F_{d}\)はNADP⁺に電子(e⁻)を渡し、FNRによって触媒(還元酵素)され、NADPHを生産する。
NADP++2e–+H⁺→NADPH
生成されたNADPHは、還元力としてカルビンサイクルにて炭素固定反応に用いられ、二酸化炭素(CO₂)に電子を与える還元力を持った物質としてブドウ糖(C₆H₁₂O₆)を作る原動力となる。
注1:電子(e⁻)を渡されたNADP⁺は負電荷を持つようになるので、ストロマ側からプロトン(H⁺)を吸収(結合)してNADPHになる。電子を与えやすい還元力を持った還元剤となる。
注2:NADPHが電子(e⁻)を失うと、水素イオン(H⁺)が分離してNADP⁺へと変わり、相手から電子を奪う酸化力を持った酸化剤となる。

「酸化とは、電子を失うこと」であり、「還元」とは、電子を得ること」である。
「酸化」と「還元」という反応は、必ず2つ同時に起きる。
ある物質が「酸化」されて電子を失えば、その電子は別の物質に受け渡され「還元」されている。
この時、相手を「酸化」する物質を「酸化剤」、同時に別の相手に「還元」する物質を「還元剤」という。
「酸化剤」は、相手から電子を奪うことで自身は「還元」され、「還元剤」は、相手に電子を与えたため自身は「酸化」されたことになる。
「酸化剤」が相手を酸化する(電子を奪う)力を「酸化力」、「還元剤」が相手を還元する(電子を与える)力を「還元力」という。
その「還元剤」から「酸化剤」へ電子が受け渡される際には、エネルギーが放出される。
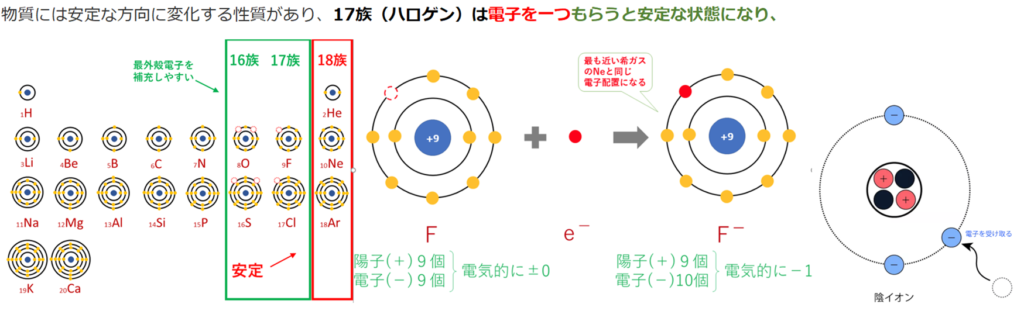
物質それぞれには、電子と本来固有の結びやすさがある。
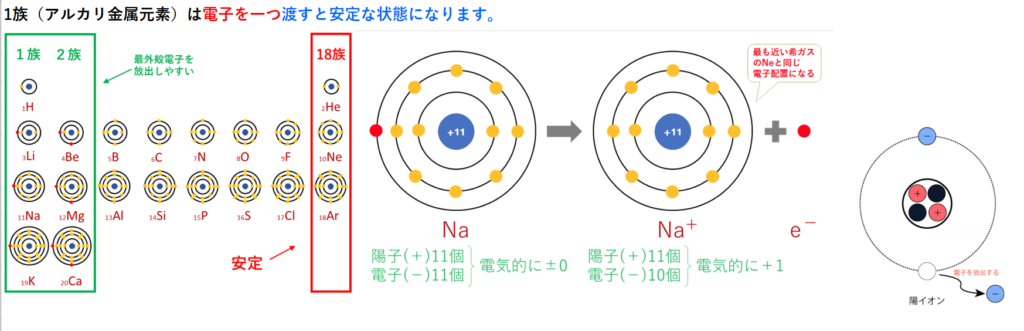
電子を与えやすい物質は、電子と結びにくい物質から、電子と結びやすい物質へと伝わっていく。
この「電子と結びやすさ」の指標を「酸化還元電位」と呼び、その差を「電位差」という。 電子の伝達にもエネルギーが放出される。「還元剤」は、「電位差」というエネルギーを、電子という形で物質内に蓄えている。
●酸化力と還元力の生成
NADPHを還元するための電子は,水の酸化により供給される。
NADPH の標準酸化還元電位は-0.32 V であり,水の酸化反応の標準酸化還元電位は+0.82 V である。
最低でもその差の 1.14 V に相当するエネルギーが外部から供給されなければ反応は進行しない。
光合成系においては,2 段階の光化学反応(PSⅡ、PSⅠ)によってこのエネルギーを供給している。
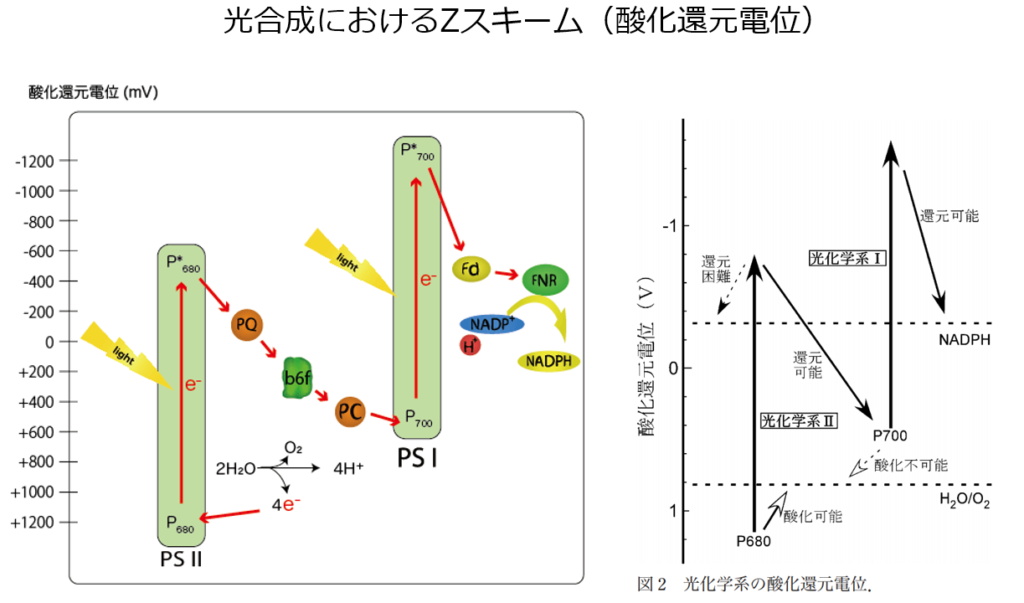
光化学系Ⅱの反応中心である P680 の酸化還元電位は、+1.1 V を超し、水を分解して電子を取り出し、酸素を発生することを可能にする(Mn₄CaO₅クラスターにて)。
励起された P680 は、数値的には直接 NADPH を還元するだけの酸化還元電位をもつが、NADPH の酸化還元電位との差は小さい。
電位差が低い為、NADPH を還元できず、NADPH を還元するために光化学系Ⅰ(P700)が必要となる。
■■『光リン酸化』■■
PSⅡの反応中心では、太陽光からのエネルギーを使って水から電子(e⁻)を取り出し、電子伝達鎖を介してPSⅠにおいてNADP⁺をNADPHに還元する。この際にプロトン(H⁺)をストロマ側から除去する。
一方、チラコイド内腔では、2分子の水分解で4つ増えたプロトン(4H⁺)と更にシトクロムb₆/fから2個のプロトン(2H⁺)が送り込まれ、水素イオン濃度が高くなり、濃度勾配が発生する。
秋田 総理,菅 倫寛,沈 建仁
岡山大学異分野基礎科学研究所・光合成・構造生物学研究コア
正味の結果は、チラコイド内腔内の低いpHとストロマ内の高いpHになる。
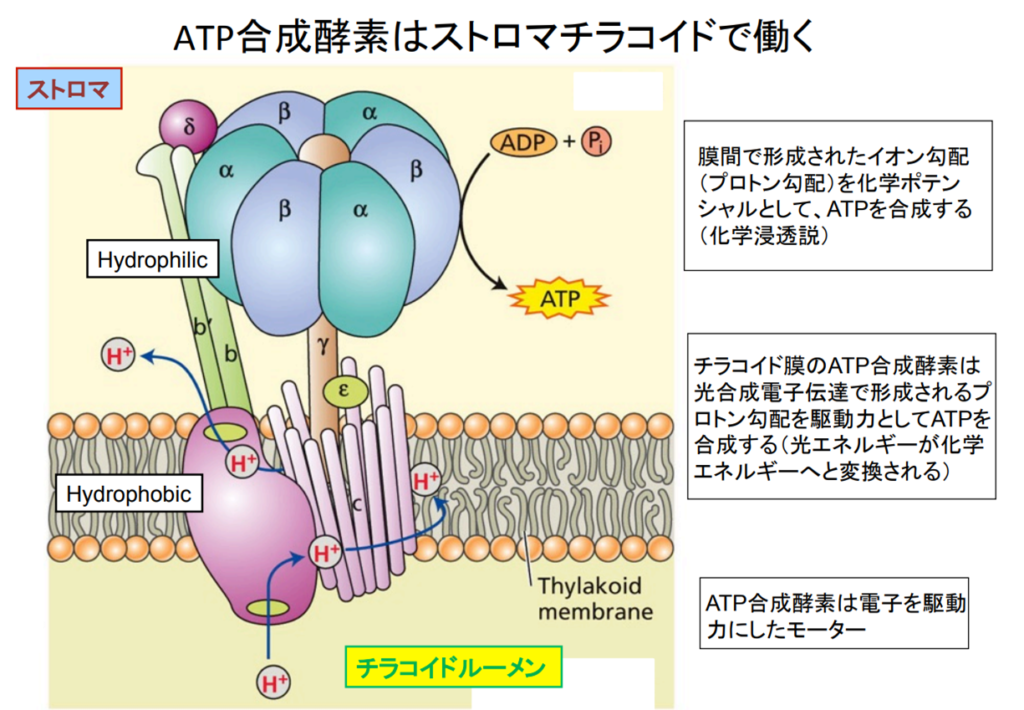
出典:4.1 生態系における光合成 4.2 地球環境と光合成 4.3 … – 岡山大学
チラコイド内腔のプロトン(H⁺)が濃度勾配によりストロマに向かって押し出される際、ATPシンターゼ(合成酵素)により摩擦熱が生み出され、この熱エネルギーで2つのリン酸を持つADPに更に活性の鋭いリン酸を結合させATPに変える。
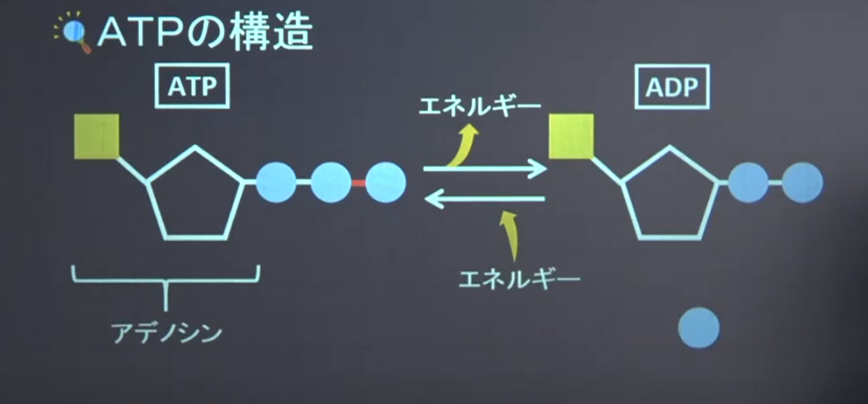
注:三リン酸の2番目と3番目に『高エネルギーリン酸結合』がしている
ATPは生体内のエネルギー貨幣とも呼ばれ、多くのエネルギー代謝に関わる事ができる大切な物質である。
ATPは二酸化炭素を糖に変える時、無リン酸を切り離す際に多量のエネルギーを放出する。
PSⅡ、PSⅠの電子伝達鎖で生成されたNADPHと光リン酸化で生成されたATPは暗反応(炭素固定反応=カルビンサイクル)にて利用される。
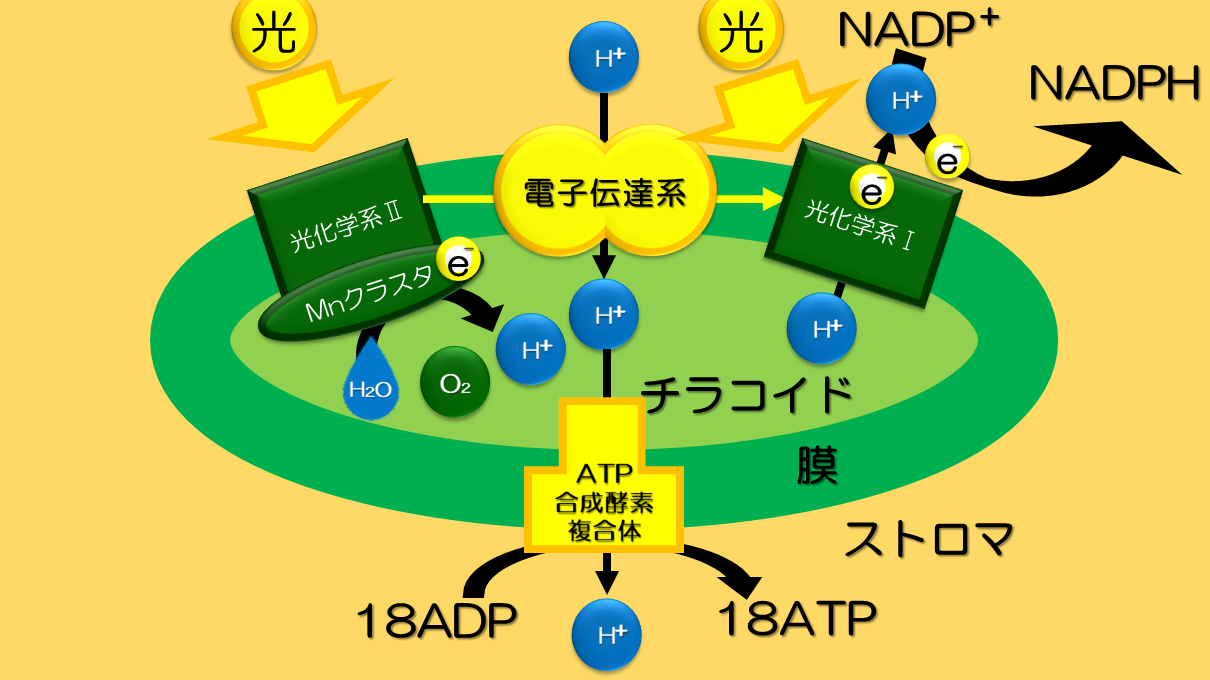
https://www.h-biology.info/high-biology/life-phenomena-and-substances/metabolism/photosynthesis/process-with-the-reaction-equation-of-photosynthesis/